

La diversité des virus de l'influenza relève principalement de deux aspects étroitement liés. D'une part, les mécanismes évolutifs principaux auxquels est soumis ce virus —la dérive et le réarrangement ou échange génétique (discutés dans un autre article de cette même rubrique)— ; de l'autre, le comportement épidémiologique du virus dans chaque espèce, lequel, comme on va le voir dans cet article, a un effet important sur les mécanismes évolutifs.
Si l'on compare à partir de la dérive génétique les virus présents chez le porc à ceux présents chez l'être humain, deux phénomènes sont principalement à souligner : 1) considérant une souche spécifique, chez l'être humain le virus varie antigéniquement (changements dans les acides aminés) plus rapidement que chez le porc, même si le taux de mutation génétique du virus est le même pour les deux espèces, et 2) globalement, la diversité du virus de l'influenza, aussi bien au niveau génétique qu'antigénique, est beaucoup plus grande chez le porc que chez l'être humain. Même si ces deux faits peuvent sembler à priori contradictoires, ils sont, comme on le verra tout de suite, parfaitement compatibles.

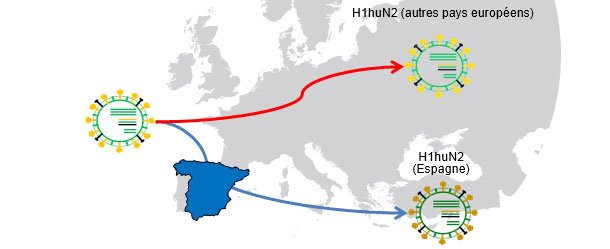
Figure 1. Ancêtre commun H1huN2 (1994) ; Isolement géographique → la dérive produit des variantes liées à chaque pays.
Dans d'autres articles de cette même section, on a vu que l'influenza peut agir endémiquement ou enzootiquement chez le porc ; autrement dit, que les virus de l'influenza peuvent "résider" dans les exploitations. Cette situation est très fréquente et signifie l'apparition de circulations récurrentes. Dans une étude de 2001 (Jong et collaborateurs) plus de 20 H1N1 porcins de type aviaire (H1avN1) ont été comparés génétiquement et antigéniquement. Ces virus avaient été isolés dans six exploitations des Pays Bas endémiquement infectées. Sur la base du gène de l'hemaglutinine (H), il a été constaté que les isolats formaient cinq groupes présentant des différences au niveau génétique et antigénique. Il faut souligner que chaque groupe correspondait à une exploitation (sauf l'un des groupes qui incluait des isolats de deux exploitations). Cette étude prouve que tous les isolats ont un H1avN1 ancêtre commun mais que, grâce à la circulation endémique, les virus évoluent et se différencient de manière isolée. Dans une autre étude (Martín-Valls et coll., 2013), réalisée dans le CReSA entre 2009 et 2012, quatre virus H1N2 ont été isolés. Lorsque l'H1 a été analysée d'un point de vue génétique, il a été observé qu'ils appartenaient tous au type H1N2 "type humain" (H1huN2), mais qu'ils formaient un groupe clairement différencié au sein des H1huN2 ; il pouvait donc en être conclu qu'il y avait eu en Espagne une évolution isolée de ce type. Ultérieurement, on a pu constater que ces virus réagissaient de manière limitée avec des sérums qui, par contre, réagissaient avec un autre H1huN2 commercial, ce qui indiquerait une certaine dérive antigénique (données non publiées). En définitive, aussi bien les facteurs épidémiologiques que l'évolution par isolement dans des exploitations en situation endémique, ou l'isolement géographique, jouent un rôle important dans l'apparition de nouvelles variantes antigéniques. Chez l'être humain, cependant, la situation est complètement différente. Dans ce cas, on peut dire que, compte tenu de la pression immunitaire globale, les virus sont forcés de changer antigéniquement pour pouvoir survivre. Actuellement, les connexions multiples et variées existant entre les pays du monde entier éliminent les situations d'isolement. Par conséquent, chez l'être humain, il existe une sélection positive globale des variantes du virus de l'influenza capables d'échapper à la réponse immunitaire, un phénomène se traduisant par un besoin constant d'une mise à jour des vaccins chez l'être humain.

Figure 2. H1huN2 est transmis dans une exploitation infectée endémiquement par H1avN1 → un réarrangement a lieu.
Le réarrangement ou échange génétique peut également être à l'origine de changements au niveau antigénique, même d'une manière plus subite. Chez l'être humain, par exemple, celle-ci a été la cause principale des trois dernières pandémies, ayant supposé l'introduction d'au moins une nouvelle hémaglutinine antigéniquement différente à la pré-existante dans la population et face à laquelle il n'y avait pas d'immunité préalable. Dans un autre article de cette rubrique on a déjà vu que, chez le porc, les virus de l'influenza porcine font l'objet de réarrangements fréquents qui, par conséquent, donnent lieu à de nouvelles souches, comme par exemple le H1avN2, un H1N2 qui croise antigéniquement avec les virus H1avN1 (Rose et coll., 2013), ou encore les H1huN1, qui croisent antigéniquement avec le H1huN2 (Martín-Valls et coll., 2013; Rose et coll., 2013). Pour que ces réarrangements se produisent, il faut que deux souches différentes infectent un même animal en même temps. Évidemment, les situations endémiques constituent des contextes idéaux pour que cela se produise. Le fait est que, lorsque des situations endémiques ont été évaluées, des évidences de réarrangement génétique ont été observées entre différentes souches et même différents sous-types (Martín-Valls et coll., 2013; Rose et coll., 2013).
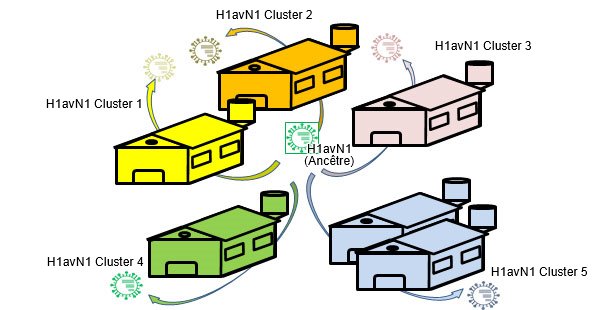
Figure 3. Les infections endémiques facilitent l'évolution isolée par dérive dans chaque exploitation → hétérogénéité génétique et antigénique par rapport à l'exploitation.
En conclusion, les virus de l'influenza porcine présentent une diversité génétique et antigénique élevée générée par les mécanismes évolutifs intrinsèques des virus. Cette diversité est renforcée par des facteurs épidémiologiques comme les situations de circulation endémique et/ou l'isolement géographique, elle est donc accentuée avec le temps. L'interaction de tous ces facteurs peut avoir des implications dans l'absence de reconnaissance immunologique ; c'est-à-dire, dans un déficit de protection croisée entre des souches appartenant à un même sous-type.

